IN: Morphogenesis: Cellular Interactions. Annals of the New York Academy of Sciences Vol. 857 pp. 268-271.
Transient membranes that are removed to complete communication between endodermal and ectodermal chambers are a feature of vertebrate morphogenesis at both ends of the embryonic axis. In each case the mechanism of opening is not completely understood.
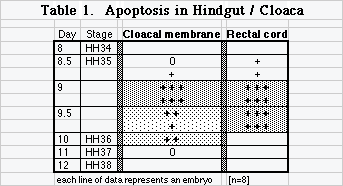
Earliest chicken cloaca is cavitated by extensive apoptosis that removes remnants of the primitive streak and the tail [6]. Subsequent cloacal morphogenesis involves cavitation of occluded cords of cells and vacuolization of membranes. "Membranes" of the hindgut are thick appositions of adjacent layers that cavitate into cords of cells and eventually open a passage or lumen. Chick cloaca develops as three successive chambers that are distinct around midgestation, but become indistinct in adults [1,4]. At day 9, two chambers, urodeum and proctodeum, are separated by cloacal membrane (a.k.a. cloacal plate, anal plate, anal membrane, urodeal membrane) whereas the coprodeum has not yet cavitated from rectal cord [10]. Between day 9 and 10 a series of cellular cords appears in cloacal plate and occluded rectal cord [5]. Cell death is not investigated in these anatomical descriptions.
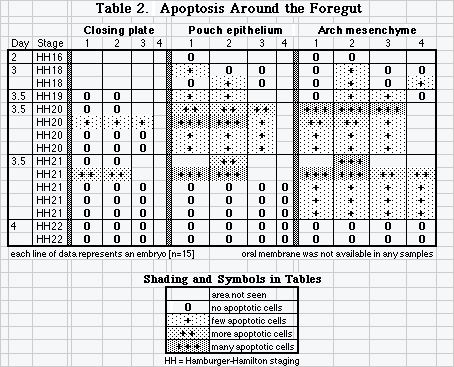
Pharyngeal membranes also fenestrate into cell cords as they rupture, and cells in pharyngeal membranes continue to proliferate during rupture [7,9]. Cell death has not been considered important in rupture of pharyngeal membranes [11], but in a preliminary survey using immunomarkers [8] we found evidence that limited cell death may have a role in rupturing pharyngeal membranes as well. Remodeling of pharyngeal pouches could also involve cell death.
Materials and Methods
White Leghorn Chicken embryos (SPAFAS) incubated at 38°C were removed at appropriate times of development, fixed in neutral-buffered formalin, and prepared with appropriate histology for applying the TUNEL method [3] (ApopTag®) to selected sections using diaminobenzidene (DAB) to mark the presence of apoptosis.
Results
Two investigators examined cloacas of 8 embryos that represented 4 days of development and Hamburger and Hamilton (HH) stages 34-38. Abundant and dense collections of apoptotic cells flanked the epithelial mass of day 9 urodeal membrane and were dispersed among emerging cell cords in cavitating rectum (Table 1).
Three investigators examined pharynx of 15 embryos representing stages 16 to 22. Apoptosis marked clearly and abundantly in locations of neural crest: head mesenchyme, arches, and around the thyroid rudiment. Apoptotic bodies were often present in foci in endodermal epithelium of pouches. Occasional apoptotic bodies marked in head ectoderm. Apoptosis was rarely localized in the pharyngeal closing plates, but one investigator found clear apoptotic markers in closing plates and shreds of ruptured plates in 2 embryos [HH 20-21, day 3.5]. No samples of oral membrane were available (Table 2).
Discussion
Extensive apoptosis removes tail gut and remnants of primitive streak [6] and apoptosis continues in a significant role in cloacal morphogenesis. What Jolly called vacuolization of epithelial masses is clear and resembles cell cords in rupturing oral membrane and septum transversum. Abundant and dense collections of apoptotic cells in day 9 urodeal/cloacal membrane and in the openings of occluded rectal cord suggest that programmed cell death is a significant contributor to vacuolization in epithelial masses in hindgut/cloaca. Positive apoptosis markers in several closing plates of 2 embryos suggest that apoptosis has at least a limited role in removing some cells of ruptured pharyngeal membrane shreds.
Apoptosis is an ephemeral process, and immunomarkers flag only late apoptosis [2]. Further investigation of more frequent time stages might reveal transient apoptosis missed by our surveys. Once detected, however, as few as 2-3% of cells marked can suggest as much as 25% of a population may be undergoing apoptosis over a day's time [2].
Apoptosis is often contemporary with cell division in a healthy embryo. It can remove cells that are no longer necessary, and it can help to limit hyperplasia. In a limited study of cell proliferation using BrdU, we found extensive proliferation in cloacal epithelium, at 10 and 11 days (unpublished observations). Further study is necessary to clarify details of chick hindgut morphogenesis.
Conclusions Apoptosis has a role in formation of cell cords and cavitations in chick embryo hindgut and cloaca, and limited apoptosis may remove some cells of pharyngeal closing plate stubs.
Acknowledgments We are grateful for financial assistance from Sigma Xi, the Howard Hughes Foundation, the Casstevens Family Fund and the Hamilton College Academic Fund for Seniors. Data reported here come from the Senior Theses of Clark, Cooney, Crary, and Payzant and summer research of Briglin.
Literature Cited
![]()
![]()
Last Modified: 3 September 1999